The 7 Viruses That Cause Human Cancers


Epstein-Barr Virus: Burkitt’s Lymphoma, Hodgkin’s Disease, and Nasopharyngeal Carcinoma
Because human tumor viruses sometimes depend on weakened host immunity, environmental factors, and host cellular mutations, one type of viruses is able to cause various types of cancer. For example, EBV is associated with Burkitt’s lymphoma, nasopharyngeal carcinoma, and some forms of Hodgkin’s disease. EBV can readily infect and alter the genetic code of human B cells, and may predispose immunosuppressed patients to malignant tumors.
Its early discovery (1964) and monoclonal genome made Epstein-Barr virus (EBV) one of the best studied examples of a cancer-causing virus. Burkitt’s lymphoma is caused by overexpression of the MYC gene, which codes for a transcription factor that modulates genes related to cell cycle progression, apoptosis, and cellular transformation. Infection by EBV may lead to a MYC mutation that allows human B cells to proliferate indefinitely. Depending on the stage of B cell development during EBV infection, EBV infection may also lead to different forms of Burkitt’s lymphoma. Recent insights indicate that the development of Burkitt’s lymphoma may be polymicrobial in nature. For example, children with recent infection by the malaria-causing Plasmodium falciparum were more likely to develop Burkitt’s lymphoma.
In patients with Hodgkin’s disease, EBV may directly trigger tumorigenesis, especially through constitutive expression of MYC. The association of EBV with Hodgkin’s disease is multifactorial, and includes factors such as country of residence, histological subtype, sex, ethnicity, and age.
Finally, EBV can cause nasopharyngeal carcinoma, which might be a result of virus latency in nasopharyngeal epithelial cells. The genetic mutations caused by EBV infection depend on the viral type and strain, but smoking has been associated with nasopharyngeal carcinoma and possibly leads to EBV reactivation.
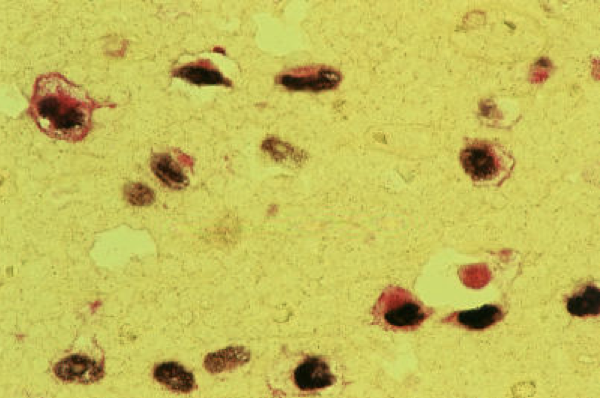
Pathologists have been able to harness recent advances in viral oncology to visualize cancer under the microscope. Double labelling of malignant cells, as in Figure 1, shows co-expression of Epstein-Barr virus early RNAs (brownish black in color) and latent membrane protein 1 (LMP1; red in color). LMP1 is a well-established surrogate marker for EBV, and is seen in Figure 1 (right) via immunohistochemistry staining of EBV infected cells.
Human Papillomaviruses 16 and 18: Cervical Carcinoma, Anal Carcinoma, Oropharyngeal Carcinoma, Penile Carcinoma
Human papillomavirus (HPV) is the most common sexually transmitted virus in the world, and is present in up to 79 million Americans. The HPV genome is divided into three regions: the noncoding upstream regulatory region; the “early” region, involved in viral replication and oncogenesis; and the “late” region, which encodes structural proteins for the viral capsid. When the HPV genome becomes integrated into human chromosomes, it disrupts an open reading frame and causes the overexpression of two viral oncogenes. These oncogenes decrease the expression of tumor suppressor protein p53 (also known as the “Guardian of the Genome”) and dysregulating the retinoblastoma protein (pRB), which can eventually lead to cancer.
In countries with accessible preventive health services, there has been a steep decline (approximately 70% from 1955 to 1992) in mortality caused by cervical cancer. This is due to the development and widespread acceptance of cervical cancer screening (Pap tests). Further, the HPV vaccination is now available for individuals up to 45 years of age, which prevents approximately 70-80% of cancer cases.
Kaposi’s Sarcoma-Associated Herpesvirus: Kaposi’s Sarcoma, Primary Effusion Lymphoma, Multicentric Castleman’s Disease
Kaposi’s sarcoma is is the most common cancer in untreated individuals with HIV positive status. Kaposi sarcoma-associated herpesvirus is also known as herpesvirus-8 (HHV-8). KSHV infects endothelial cells and modulates pathways that control cell proliferation, gene expression, and metabolism. KSHV may selectively activate and suppress its lytic replication cycles during co-infections with other microbes by sensing resources and cellular conditions. In Figure 2 (left), one can visualize Kaposi sarcoma, caused by KSHV, on the lungs of a patient with AIDS. Infection by multiple viruses is likely to be synergistic, an important consideration for immunocompromised individuals because they are more likely to develop infections.
Hepatitis B Virus and Hepatitis C Virus: Hepatocellular Carcinoma

Hepatitis B virus (HBV) is an enveloped DNA hepadnavirus that can cause hepatocellular carcinoma (HCC). HCC is the third leading cause of cancer deaths worldwide. HBV can integrate itself into the human genome and replicate within liver cells, directly increasing carcinogenic activity through several signaling pathways in the liver. Hepatitis B surface antigen (HBsAg), a popular marker for the medical diagnosis of active hepatitis B, accumulates within the endoplasmic reticulum inducing oxidative stress to cause liver damage. HBsAg also increases cell motility and decreases apoptosis.
At least two viral proteins (including HBV X protein and Hepatitis B surface antigen) and several human mRNAs (such as miR-122) have been linked to hepatocellular carcinoma. HBV X protein is found at high cytosolic concentrations in HBV-infected hepatocytes. HBV X protein has a complex role in hepatocellular carcinoma. It has been shown to decrease apoptosis induced by several cell mediators, such as p53, Fas, TNF, and TGF-beta, but it has also been shown to increase apoptosis by promoting the production of reactive oxygen species (ROS) that damage hepatocytes.
Hepatitis C virus (HCV), an RNA virus, is the most common cause of hepatocellular carcinoma. Like the Hepatitis B virus, HCV has a multifactorial role in the development of liver cancer. For example, HCV can activate pathways that lead to liver fibrosis, impact apoptosis and cellular survival, interact with immune cells, and affect metabolic processes. In contrast with HBV, HCV cannot integrate into the human genome. The progression of HCV can take up to 20 to 40 years, and involves progressive liver fibrosis through irreversible genetic and epigenetic alterations. The malignant transformation of liver cells in HCC involves a variety of pathways, and may cause mutations of genes such as p53, PIKCA, and beta-catenin. Liver cancer risk appears to be dependent on viral genotype, and HCV genotype 3, 25, 26, and 27 are more carcinogenic.
Human Adult T-cell Leukemia Virus Type 1 (HTLV-1): T-cell Leukemia
HTLV-1 is a retroviral infection that affects white blood cells known as T cells. Although HTLV-1 infection rarely causes serious disease, it may lead to adult T-cell leukemia in 2-5% of infected individuals or HTLV-1-associated muscle disorders in 0.25-2% of infected people. Despite the association of HTLV-1 with cancer, its cancer-causing mechanisms are poorly understood due to difficulties propagating this virus in tissue culture. However, T cells that are infected by HTLV-1 can undergo genetic and epigenetic alterations. Additionally, Gag polyprotein may be a major player because it is responsible for the assembly and release of viral particles. In Figure 3 (right), HTLV-1 is visible via cryogenic transmission electron microscopy. Through cryogenic TEM, the lattice-like structure of Gag proteins can be studied and compared to other retroviruses (such as HIV).
Merkel Cell Polyomavirus: Merkel Cell Carcinoma

The final virus linked to human cancers is the Merkel cell polyomavirus (MCV). A rare skin cancer, Merkel cell carcinoma (MCC) was the first human cancer to be associated with a polyomavirus. To date, it is the only polyomavirus with a robust collection of scientific evidence supporting its classification as a causative agent of a human malignancy. MCC is considered a neuroendocrine cancer that commonly occurs in sun-exposed individuals with weakened immune systems (for example, in the elderly). Some subtypes of MCV are residents of the skin microbiome, but studies of the MCV viral life cycle remain inconclusive.
This article is the second of a 2-part series on cancer virology. An introduction to the infectious causes of cancer can be found here.
© American Society for Microbiology | Author: Jennifer Brubaker, Ohio University Heritage College of Osteopathic Medicine